Os músculos esqueléticos são formados por feixes paralelos de numerosas células muito longas, as fibras musculares, cuja membrana plasmática é chamada sarcolema (sarkos, em grego, significa carne). O citoplasma (sarcoplasma) destas células contém centenas de filamentos contráteis organiza dos em paralelo, as miofibrilas. Estas são compostas por dois tipos principais de filamentos protéicos, filamentos grossos e filamentos finos, e sua disposição resulta no padrão de bandas de diferentes refringências, que conferem uma aparência estriada a fibra muscular (ao microscópio óptico). Ao microscópio eletrônico, é possível visualizar zonas claras e escuras alternadas: as bandas I(de isotrópicas), contendo somente filamentos finos, e as bandas A (de anisotrópicas), contendo filamentos grossos e finos parcialmente sobrepostos. No centro da banda I aparece a linha Z (ou disco Z). Os filamentos finos se interdigitam nos filamentos grossos. Os arranjos de filamentos grossos e finos formam o sistema contrátil, e a unidade repetitiva dos arranjos de filamentos grossos e delgados, em cada miofibrila, é a unidade básica contrátil, chamada de sarcômero, região compreendida entre duas linhas Z.
dos em paralelo, as miofibrilas. Estas são compostas por dois tipos principais de filamentos protéicos, filamentos grossos e filamentos finos, e sua disposição resulta no padrão de bandas de diferentes refringências, que conferem uma aparência estriada a fibra muscular (ao microscópio óptico). Ao microscópio eletrônico, é possível visualizar zonas claras e escuras alternadas: as bandas I(de isotrópicas), contendo somente filamentos finos, e as bandas A (de anisotrópicas), contendo filamentos grossos e finos parcialmente sobrepostos. No centro da banda I aparece a linha Z (ou disco Z). Os filamentos finos se interdigitam nos filamentos grossos. Os arranjos de filamentos grossos e finos formam o sistema contrátil, e a unidade repetitiva dos arranjos de filamentos grossos e delgados, em cada miofibrila, é a unidade básica contrátil, chamada de sarcômero, região compreendida entre duas linhas Z.
Filamentos grossos: Miosina
Os filamentos grossos são constituídos basicamente por miosina, uma proteína com seis cadeias polipeptídicas: um par de cadeias grandes e pesadas, e dois pares de cadeias leves. As cadeias se organizam em uma porção fibrosa, a cauda da miosina, e duas porções globulares idênticas, as cabeças da miosina. A cauda é um longo bastão em dupla hélice, resultante da associação das duas cadeias pesadas, com estrutura em α-hélice, que se espiralam uma em torno da outra, ao longo de grande extensão da região carboxila terminal, formando uma super-hélice. Na extremidade amino terminal da dupla hélice, as cadeias pesadas dobram-se separadamente e cada uma se une a duas cadeias leves formando as cabeças da miosina. Cada cabeça possui um domínio catalítico e um domínio mecânico (ou regulador). O domínio catalítico contém o centro ativo com atividade ATPásica e o sítio de ligação dos filamentos finos. O domínio mecânico é constituído por um trecho em hélice da cadeia pesada ao qual se entrelaçam as cadeias leves; situa-se entre o domínio catalítico e a cauda da miosina, aos quais se une por ligações flexíveis. As cabeças da miosina também são chamadas de S1, por corresponderem a um dos fragmentos peptídicos produzidos por digestão parcial da miosina com enzimas proteolíticas, o fragmento S1(de subfragment 1). Os filamentos grossos são formados por centenas de moléculas de miosina, associadas cauda a cauda. As cabeças ficam orientadas em direções opostas em relação ao centro do sarcômero, onde se encontram somente segmentos lineares, as caudas. As cabeças projetam-se a intervalos regulares, podendo ligar-se aos filamentos finos, constituindo as pontes cruzadas. A porção central dos filamentos grosso não possui pontes cruzadas, e estas se projetam em grupos de três a partir do filamento grosso. Os filamentos grossos no músculo estriado têm 1,6 µm de comprimento e contém uma estimativa de 300 a 400 pontes cruzadas.

Filamentos finos: Actina, tropomiosina e troponina
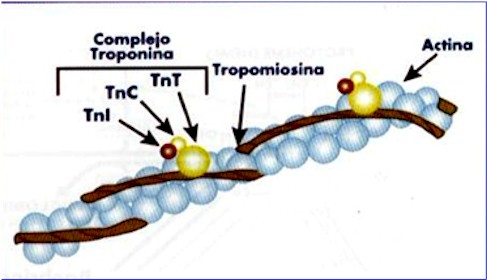
 Cada filamento fino contém uma molécula de actina F associada a duas outras proteínas, a tropomiosina e a troponina. A tropomiosina é uma molécula em forma de bastão, composta por duas cadeias polipeptídicas separadas. Os polipeptídios individuais têm uma estrutura básica em α-helice, e os dois peptídeos helicoidais estão enrolados, um ao redor do outro, para formar uma super-hélice; ela se insere ao longo do sulco entre as duplas hélices da actina F, cobrindo sete monômeros de actina. A união de varias moléculas de tropomiosina constitui um filamento que se estende por toda a superfície da actina F. Cada molécula de tropomiosina esta unida a uma molécula de troponina. Esta é a proteína reguladora da interação da actina com o filamento grosso. A troponina é uma proteína composta por três subunidades: a troponina I (TnI), que se une a actina inibindo a ligação de S1 à actina; a troponinaT (TnT), que se une a tropomiosina; e a troponina C (TnC), que pode se unir a íons de cálcio.
Cada filamento fino contém uma molécula de actina F associada a duas outras proteínas, a tropomiosina e a troponina. A tropomiosina é uma molécula em forma de bastão, composta por duas cadeias polipeptídicas separadas. Os polipeptídios individuais têm uma estrutura básica em α-helice, e os dois peptídeos helicoidais estão enrolados, um ao redor do outro, para formar uma super-hélice; ela se insere ao longo do sulco entre as duplas hélices da actina F, cobrindo sete monômeros de actina. A união de varias moléculas de tropomiosina constitui um filamento que se estende por toda a superfície da actina F. Cada molécula de tropomiosina esta unida a uma molécula de troponina. Esta é a proteína reguladora da interação da actina com o filamento grosso. A troponina é uma proteína composta por três subunidades: a troponina I (TnI), que se une a actina inibindo a ligação de S1 à actina; a troponinaT (TnT), que se une a tropomiosina; e a troponina C (TnC), que pode se unir a íons de cálcio.
*Bibliografia:
http://www.ufmt.br/bionet/principal.htm http://www.portalesmedicos.com/images/publicaciones/0907_marcadores_bioquimicos_cardiacos/marcadores_cardiacos_troponina_2.jpg
MARZZOCO, A.; TORRES, B.B. Bioquímica básica. 3ª edição. Ed. Guanabara Koogan, Rio de Janeiro, 2007.
BERNE, RM; LEVY, MN. Fisiologia. 4ª edição, Ed. Guanabara Koogan, Rio de Janeiro, 2000
Postado por: Bruna Neuhauss

{kind=link}